Es un tipo de globulina.
Que se denomina así por aparecer en último lugar cuando se separan las proteínas del suero sanguineo por medio de una electroforesis.
El término gamaglobulina es usado frecuentemente como sinónimo de inmunoglobulina, debido a que la mayoría de las inmunoglobulinas son gamma globulinas; sin embargo, dado que algunas inmunoglobulinas migran a las regiones beta o alfa, el término tiende a usarse menos.
Las inmunoglobulinas son proteínas heterodiméricas compuestas por dos cadenas pesadas (H) y dos cadenas ligeras (L). Se pueden separar funcionalmente en dominios variables (V) que se unen a antígenos y dominios constantes (C) que especifican funciones efectoras tales como activación del complemento o unión a receptores Fc.
Los dominios variables se crean por medio de una serie compleja de eventos de reordenamiento génico, y luego pueden someterse a hipermutación somática después de la exposición al antígeno para permitir la maduración de afinidad.
Cada dominio V se puede dividir en tres regiones de variabilidad de secuencia, denominadas regiones determinantes de complementariedad, o CDR, y cuatro regiones de secuencia relativamente constante denominadas regiones marco, o FR.
Las tres CDR de la cadena H se combinan con las tres CDR de la cadena L para formar el sitio de unión al antígeno, como se define clásicamente. Hay cinco clases principales de dominios C de cadena pesada.
Cada clase define los isotipos IgM, IgG, IgA, IgD e IgE. IgG se puede dividir en cuatro subclases, IgG1, IgG2, IgG3 e IgG4, cada una con sus propias propiedades biológicas; e IgA pueden dividirse de manera similar en IgA1 e IgA2.
Los dominios constantes de la cadena H se pueden cambiar para permitir la función efectora alterada mientras se mantiene la especificidad del antígeno.
Más de 100 años de investigación sobre la estructura y función de la gamaglobulina solo han servido para enfatizar la naturaleza compleja de esta proteína. Típicamente, los receptores se unen a un conjunto limitado y definido.
Sin embargo, aunque la gamaglobulina individual también se une a un conjunto limitado y definido de ligandos, las inmunoglobulinas como población pueden unirse a una matriz virtualmente ilimitada de antígenos que comparten poca o ninguna similitud.
Esta propiedad de unión ajustable depende de una compleja serie de mecanismos que alteran el ADN de las células B individuales.
La gamaglobulina también sirve para dos propósitos:
- Receptores de superficie celular para el antígeno que permite la señalización celular.
- La activación celular y el de las moléculas efectoras solubles que pueden unir y neutralizar antígenos individualmente a distancia.
Estructura de la gamaglobulina
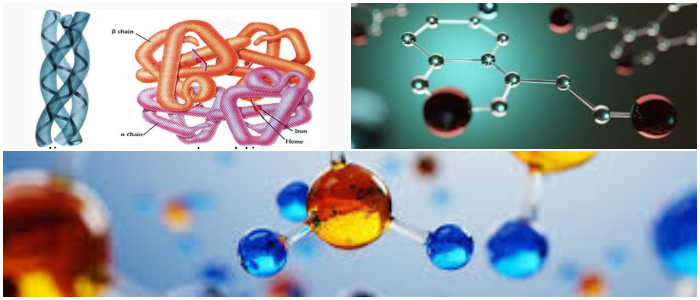
Las moléculas de anticuerpos (o gamaglobulinas) son glicoproteínas compuestas de una o más unidades, cada una con cuatro cadenas de polipéptidos; dos cadenas pesadas idénticas (H) y dos cadenas ligeras idénticas (L).
Los extremos amino terminales de las cadenas polipeptídicas muestran una variación considerable en la composición de aminoácidos y se denominan regiones variables (V) para distinguirlas de las regiones relativamente constantes (C).
Cada cadena L consta de un dominio variable, VL y un dominio constante, CL. Las cadenas H constan de un dominio variable, VH y tres dominios constantes CH1, CH2 y CH3.
Cada cadena pesada tiene aproximadamente el doble de aminoácidos y peso molecular (~ 50,000) que cada cadena ligera (~ 25,000), dando como resultado un peso molecular del monómero de inmunoglobulina total de aproximadamente 150,000.
Las cadenas pesadas y ligeras se mantienen juntas por una combinación de interacciones no covalentes y enlaces disulfuro intercatenarios covalentes, formando una estructura simétrica bilateral.
Las regiones V de las cadenas H y L comprenden los sitios de unión a antígeno de las moléculas de inmunoglobulina (Ig). Cada monómero de Ig contiene dos sitios de unión a antígeno y se dice que es bivalente.
La región de bisagra es el área de las cadenas H entre el primer y el segundo dominio de la región C y se mantiene unida mediante enlaces disulfuro. Esta región de bisagra flexible (que se encuentra en IgG, IgA e IgD, pero no IgM o IgE) permite que varíe la distancia entre los dos sitios de unión a antígeno.
Clases de gamaglobulinas
Las cinco clases principales de gamaglobulina son IgG, IgM, IgA, IgD e IgE. Estos se distinguen por el tipo de cadena pesada que se encuentra en la molécula.
Las moléculas IgG tienen cadenas pesadas conocidas como cadenas gamma; Las IgM tienen cadenas mu; Las IgA tienen cadenas alfa; Las IgE tienen cadenas epsilon; e IgD tienen cadenas delta.
Las diferencias en los polipéptidos de cadena pesada permiten que estas gamaglobulinas funcionen en diferentes tipos de respuestas inmunes y en etapas particulares de la respuesta inmune.
Las secuencias de proteínas polipeptídicas responsables de estas diferencias se encuentran principalmente en el fragmento Fc. Si bien hay cinco tipos diferentes de cadenas pesadas, solo hay dos tipos principales de cadenas ligeras: kappa (κ) y lambda (λ).
Las clases de anticuerpos difieren en la valencia como resultado de diferentes números de unidades similares a Y (monómeros) que se unen para formar la proteína completa.
Por ejemplo, en humanos, los anticuerpos IgM en funcionamiento tienen cinco unidades en forma de Y (pentámero) que contienen un total de 10 cadenas ligeras, 10 cadenas pesadas y 10 de unión al antígeno.
Subclases de gamaglobulinas
Además de las principales clases de gamaglobulinas, existen varias subclases de Ig en todos los miembros de una especie animal particular.
Los anticuerpos se clasifican en subclases según las diferencias menores en el tipo de cadena pesada de cada clase de Ig. En humanos hay cuatro subclases de IgG: IgG1, IgG2, IgG3 e IgG4 (numeradas en orden de concentración decreciente en suero).
La varianza entre las diferentes subclases es menor que la varianza entre las diferentes clases. Por ejemplo, IgG1 está más estrechamente relacionado con IgG2, IgG3 e IgG4 que con IgA, IgM, IgD o IgE.
En consecuencia, las proteínas de unión a anticuerpos (p. Ej., Proteína A o Proteína G) y la mayoría de los anticuerpos secundarios utilizados en los métodos de inmunodetección reaccionan de forma cruzada con múltiples subclases pero generalmente no con múltiples clases de Ig.
Anticuerpos policlonales y monoclonales
Los anticuerpos (cualquiera que sea su clase o subclase) se producen y purifican en dos formas básicas para su uso como reactivos en inmunoensayos: policlonales y monoclonales.
Típicamente, la respuesta inmunológica a un antígeno es heterogénea, dando como resultado muchas líneas celulares diferentes de linfocitos B (precursores de células plasmáticas) que producen anticuerpos contra el mismo antígeno.
Todas estas células se originan a partir de células madre comunes, pero cada una desarrolla la capacidad individual de producir un anticuerpo que reconoce un determinante particular (epítopo) en el mismo antígeno.
Como consecuencia de esta respuesta heterogénea, el suero de un animal inmunizado contendrá numerosos clones de anticuerpos específicos de antígenos, potencialmente de varias clases diferentes de gamaglobulinas y subclases que comprenden generalmente de 2 a 5% de la inmunoglobulina total.
Debido a que contiene esta colección heterogénea de gamaglobulinas de unión a antígeno, un anticuerpo purificado a partir de dicha muestra se denomina anticuerpo policlonal.
Los anticuerpos policlonales, que generalmente se purifican directamente del suero, son especialmente útiles como anticuerpos secundarios marcados en inmunoensayos.