Es una molécula biológica muy importante.
Está involucrado en una serie de procesos biológicos y es esencial en la respiración celular.
Se encuentra comúnmente como uno de los productos finales de la glucólisis, que luego se transporta a las mitocondrias para participar en el ciclo del ácido cítrico. En ausencia de oxígeno, o cuando la demanda de oxígeno supera al suministro, el piruvato puede someterse a fermentación para producir lactato.
Tanto el piruvato como el lactato pueden usarse para regenerar la glucosa.
Hay una creciente evidencia de que puede influir directamente en la actividad nuclear y las modificaciones epigenéticas, formando la interfaz entre el genoma y el estado metabólico de la célula.
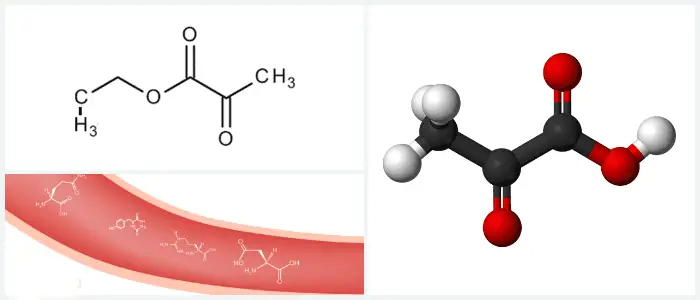
También llamado ácido pirúvico, es una molécula orgánica y consiste en una cadena principal de 3 carbonos.
Funciones del piruvato
El piruvato es necesario en no menos de seis vías metabólicas. Está involucrado en las vías de síntesis y descomposición.
La función primaria de la molécula es actuar como la molécula de transporte que lleva los átomos de carbono a las mitocondrias para la oxidación completa en dióxido de carbono.
Al final de la glucólisis en el citoplasma, las moléculas de piruvato generadas a partir de la glucosa se transportan a la matriz de la mitocondria a través de dos proteínas: portadores Piruvato Mitocondriales 1 y 2 (MPC1, MPC2).
Dentro de la matriz de las mitocondrias, un importante complejo multienzimático llamado complejo piruvato deshidrogenasa (PDC) cataliza reacciones de descarboxilación y oxidación con el fin de generar acetil coenzima A (acetil-CoA).
La primera enzima en este complejo se llama piruvato deshidrogenasa y elimina el grupo ácido carboxílico (descarboxilatos) de la molécula.
El resultado de esta reacción deja una molécula de dos carbonos que contiene un grupo metilo y un grupo carbonilo.
La segunda y tercera enzimas de PDC luego oxidan el carbono carbonilo y catalizan un enlace covalente a CoA a través de un enlace tioéster. Este enlace tioéster se puede hidrolizar con la liberación de energía.
De hecho, la hidrólisis de este enlace libera más energía que la conversión de ATP a ADP y, por lo tanto, también proporciona el ímpetu inicial para los primeros pasos del ciclo del ácido cítrico.
Recientemente, se ha llamado la atención sobre el papel de las moléculas de piruvato para influir en la acetilación de las moléculas de histonas en todo el genoma.
La acetilación de histonas es una modificación epigenética que puede cambiar la actividad transcripcional general de la célula e influir en el ciclo celular y la mitosis.
Esta modificación de histona requiere la presencia de acetil-CoA. La acetil-CoA también se genera a través de PDC en el núcleo, a través del transporte de todo el complejo enzimático desde la mitocondria hasta el núcleo.
La concentración de este complejo en el núcleo depende del ciclo celular, el entorno externo y la disponibilidad de factores de crecimiento y nutrientes. En ausencia de suficiente actividad de PDC, la transición de la célula hacia la mitosis (específicamente, la síntesis de ADN durante la fase S) se ve obstaculizada.
Otra enzima asociada con el metabolismo del piruvato que también está presente en el núcleo es la piruvato quinasa, la enzima involucrada en la última reacción de la glucólisis, que genera piruvato a partir de PEP.
Esta quinasa desempeña un papel interesante dentro del núcleo, fosforilando proteínas nucleares utilizando la PEP como donador de fosfato.
Esto, a su vez, conduce a la generación de piruvato, que puede ser utilizado por el PDC para crear acetil-CoA.
La fosforilación de residuos clave en histonas también mejora su acetilación, una vez más un paso crucial en la progresión de la célula de la fase G1 a S del ciclo celular.
Si la respiración aeróbica no es posible, el piruvato puede fermentarse a lactato en el citoplasma para generar NADH y así aumentar la disponibilidad de ATP para la célula.
La enzima involucrada en la fermentación de piruvato también puede catalizar la reacción inversa, formando piruvato a partir de lactato.
Esto es particularmente importante en las células del hígado, donde este es un proceso esencial durante el período de recuperación después del ejercicio.
Además, el piruvato funciona como uno de los puntos de partida para la gluconeogénesis, lo que permite que la célula genere glucosa a partir de fuentes que no sean carbohidratos.
Este proceso es importante para el funcionamiento del cerebro durante el ayuno, ya que los tejidos en el cerebro usan la glucosa como fuente primaria de energía.
El piruvato también está implicado en la generación de aminoácidos no esenciales, así como en muchas rutas bioquímicas que implican el metabolismo de los lípidos.
El piruvato y la respiración celular
Las moléculas de piruvato se forman durante una serie de reacciones importantes llamadas glucólisis.
La glucólisis es la vía de descomposición de las moléculas de glucosa y el primer paso de la respiración celular. La glucólisis es completamente opuesta a la gluoconogénesis porque una molécula de 6 carbonos se descompone en dos compuestos de 3 carbonos.
Es extremadamente importante para los mamíferos porque es el comienzo de nuestro ciclo energético.
Cuando comemos carbohidratos, como galletas, papas y pan, nuestros cuerpos absorben glucosa en el torrente sanguíneo.
Una vez que la glucosa se descompone, se forman moléculas de piruvato. Estas moléculas continúan produciendo más energía para las células.
Estructura
Esta molécula es la base conjugada del ácido pirúvico, una molécula de tres carbonos que contiene un grupo ácido carboxílico y un grupo funcional cetona.
La fórmula química para el ácido pirúvico es C 3 H 4 O 3 y para su forma desprotonada es C 3 H 3 O 3.
El átomo de carbono que forma el ácido carboxílico a menudo se denomina el primer átomo de carbono, y el número aumenta a lo largo de la cadena principal de carbono, lejos del extremo del ácido carboxílico.
En el piruvato, el grupo cetona está unido al segundo átomo de carbono, también conocido como carbono α, ya que está más cerca del grupo funcional principal; el tercer carbono comprende un grupo metilo.
Es, por lo tanto, el α-cetoácido más simple y de acuerdo con la nomenclatura oficial de IUPAC, se llama ácido α-ceto propanoico.
Contiene tres átomos que pueden actuar como donantes de enlaces de hidrógeno y un átomo que puede ser un aceptor de enlaces de hidrógeno.
Al igual que otros cetoácidos, el ácido pirúvico también puede tautomerizar desde su forma de cetona a su forma enólica, que contiene un doble enlace y un alcohol.
Esto es particularmente importante en el último paso de la glucólisis.
Otros α-cetoácidos implicados en la respiración celular incluyen ácido oxaloacético, ácido α-cetoglutárico y ácido oxalosuccínico.
Generación de piruvato
El piruvato se genera por dos métodos principales, a través de la vía glucolítica y a través del metabolismo de los aminoácidos.
Mientras que las proteínas suministran casi el 10% de las necesidades de energía del cuerpo, solo algunos aminoácidos se canalizan a través del piruvato hacia la maquinaria respiratoria celular.
Los que lo hacen se clasifican como aminoácidos glucogénicos, mientras que otros que generan acetil-CoA o acetoacetato se clasifican como aminoácidos cetogénicos.
El lactato producido por fermentación anaeróbica también puede regenerar piruvato, especialmente a través de la actividad de las enzimas en el hígado. Otras fuentes menores incluyen los intermedios del ciclo del ácido cítrico.
Glicólisis
La glucólisis comienza con el monosacárido de seis carbonos – glucosa. En los primeros pasos de esta vía bioquímica, la glucosa sufre fosforilación e isomerización para producir fructosa-6-fosfato.
Otra reacción de fosforilación facilita la división de este azúcar hexosa en dos moléculas de 3 carbonos: gliceraldehye phosphate (G3P) y dihydroxy acetone phosphate (DHAP).
Estos pasos iniciales requieren la entrada de energía y utilizan dos moléculas de ATP por cada molécula de glucosa, pero dan como resultado la transformación principal de una hexosa en dos moléculas de triosa.
A partir de entonces, G3P se convierte en ácido pirúvico, que existe como su base conjugada a concentración fisiológica y pH.
Este proceso ocurre a través de un conjunto de cinco reacciones bioquímicas, liberando dos moléculas de ATP y una molécula de NADH para cada molécula de G3P.
La penúltima molécula en esta cadena de reacciones se llama fosfoenol piruvato (PEP). PEP es el éster fosforilado del piruvato en su forma enólica isomérica.
El PEP pierde un grupo fosfato para generar piruvato y el resto de fosfato liberado se transfiere a ADP, formando ATP.
Esta reacción es catalizada por una enzima llamada piruvato cinasa (PK).
La reacción forma uno de los pasos limitantes de la velocidad de la glucólisis que puede determinar la velocidad de reacción global, ya que es una de las reacciones más lentas en la cadena.
Para todos los propósitos prácticos, es irreversible, a diferencia de la mayoría de las reacciones catalizadas por enzimas, especialmente debido a que el piruvato a menudo se traslada rápidamente a las mitocondrias o fermenta para formar lactato.
Cuando la glucosa necesita ser generada a partir de fuentes que no sean carbohidratos (gluconeogénesis), por ejemplo, las concentraciones cambiantes de reactivos y productos no inducen PK para catalizar la reacción inversa, formando PEP a partir de piruvato.
De hecho, durante la gluconeogénesis, la PK se desactiva mediante la fosforilación y la PEP se desvía hacia una cascada de reacciones diferente.
Metabolismo de aminoácidos
Seis aminoácidos principales se pueden metabolizar para producir piruvato: alanina, cisteína, serina, glicina, treonina y triptófano.
De estos, la alanina y la serina tienen tres átomos de carbono y, por lo tanto, son los más fáciles de transformar. Estas reacciones implican una sola enzima que cataliza esencialmente la sustitución del grupo funcional amina por una cetona.
Las enzimas también se llaman transaminasas por esta razón. La cisteína, aunque también contiene tres átomos de carbono, tiene que someterse a un paso adicional para eliminar el átomo de azufre.
La glicina, por otro lado, tiene solo dos átomos de carbono. Por lo tanto, primero se convierte en un aminoácido de tres carbonos – serina – antes de someterse a la desaminación.
La acción de la enzima serina deshidratasa cataliza su conversión en piruvato.
En una estrategia similar, tres grupos alquilo de triptófano se convierten primero a alanina antes de transformarse en una molécula de piruvato mediante la acción de la enzima alanina transaminasa.
La treonina sigue un camino aún más largo, primero se convierte en glicina y luego en serina antes de ser actuada por la serina deshidratasa.