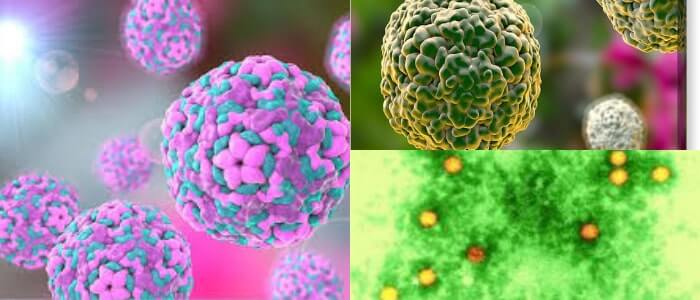
Es una familia de virus pequeños, los cuales son icosaédricos y con muy diversos genomas de ARN de cadena positiva de una sola hebra.
Las características de todos los miembros de la familia son tres proteínas de la cápside con plegamiento en barril b, procesamiento de poliproteínas por la cisteína proteinasa codificada por el virus y replicación por una ARN polimerasa dependiente de ARN con motivo de secuencia YGDD.
La familia comprende 35 géneros que contienen 80 especies, pero muchos virus están actualmente en espera de clasificación.
Los picornavirus pueden causar infecciones subclínicas en humanos y animales, o enfermedades que van desde una enfermedad febril leve hasta enfermedades graves del corazón, el hígado y el sistema nervioso central.
¿Qué es un picornavirus?
Un picornavirus es un virus perteneciente a la familia Picornaviridae. Los picornavirus son virus de ARN de cadena positiva no envueltos con una cápside icosaédrica.
El ARN genómico es inusual porque tiene una proteína en el extremo 5 ‘que se utiliza como un cebador para la transcripción por ARN polimerasa.
El nombre se deriva del pico, que significa pequeño, y el ARN, que se refiere al genoma del ácido ribonucleico, por lo que «pico-rna-virus» literalmente significa pequeño virus ARN. Los picornavirus están separados en varios géneros e incluyen muchos patógenos importantes de humanos y animales.
Las enfermedades que causan son variadas, que van desde agudas «frío común», como las enfermedades, a la polio, a las infecciones crónicas del ganado. Se siguen describiendo especies adicionales que no pertenecen a ninguno de los géneros reconocidos.
Familia de virus del picornaviridae:
- Ampivirus.
- Aftovirus.
- Aquamavirus.
- Avihepatovirus.
- Avisivirus.
- Cardiovirus.
- Cosavirus.
- Dicipivirus.
- Enterovirus.
- Erbovirus.
- Gallivirus.
- Harkavirus.
- Hepatovirus.
- Hunnivirus.
- Kobuvirus.
- Kunsagivirus.
- Limnipivirus.
- Megrivirus.
- Mischivirus.
- Mosavirus.
- Oscivirus.
- Parechovirus.
- Pasivirus.
- Passerivirus.
- Potamipivirus.
- Rabovirus.
- Rosavirus.
- Sakobuvirus.
- Salivirus.
- Sapelovirus.
- Senecavirus.
- Sicinivirus.
- Teschovirus.
- Torchivirus.
- Tremovirus.
Morfología de un virión
Los viriones consisten en una cápside, sin envoltura, que rodea un núcleo de ssRNA. Las estructuras cristalinas disponibles indican que las partículas tienen un diámetro de 30-32 nm.
Las micrografías electrónicas no revelan proyecciones sobre los viriones de la mayoría de los picornavirus, la partícula del virus aparece como una esfera casi sin rasgos distintivos; sin embargo, los kobuvirus muestran una estructura de superficie que es distinta de los pequeños virus de estructura redonda (astrovirus y calicivirus).
La cápside está compuesta por 60 unidades idénticas (protómeros). Los picornavirus con cuatro proteínas de la cápside tienen tres proteínas de superficie, 1B, 1C y 1D, de 24-41 kDa, y una proteína interna, 1A de 5.5-13.5 kDa; sin embargo, muchos picornavirus tienen tres proteínas de la cápside ya que 1AB (VP0) permanece sin escindir.
El protómero total es 80–97 kDa. Las proteínas 1A, 1B, 1C y 1D también suelen denominarse VP4, VP2, VP3 y VP1, respectivamente.
Las proteínas 1B, 1C, 1D y la 1AB no escindida poseen cada una una estructura central que comprende un sándwich b de ocho cadenas («barril b»).
Las secuencias de proteínas de la cápside (CP) en muchos géneros revelan similitudes con las de la superfamilia de proteínas «tipo rhv», y pueden contener un «sitio de unión al inhibidor» conservado que, en el caso de algunos rinocerontes y enterovirus, se ha demostrado que Se unen antivirales activos.
Los barriles B se empaquetan juntos en la cápside con T = 1, pseudo T = 3, simetría icosaédrica.
Estas características estructurales son compartidas por todos los miembros de la orden Picornavirales con estructuras atómicas resueltas.
Un ejemplo es el Virus de parálisis del grillo ( Dicistroviridae ), virus infeccioso flacherie ( Iflaviridae ) y los comovirus (virus del mosaico de caupí, virus del moteado de la vaina de frijol y virus del moteado del trébol rojo).
Los géneros difieren en los bucles externos que interconectan las hebras b. Estos bucles explican las diferencias en el alivio de la superficie de cada género y en el grosor de la pared de la cápside. El ensamblaje se produce a través de intermedios pentaméricos (pentámero: cinco protómeros).
Las proteínas dentro de cada pentámero se mantienen juntas por una red interna formada por los extremos N de los tres principales PC, los extremos C que se encuentran en la superficie externa de la cápside.
Propiedades fisicoquímicas y físicas
El peso molecular de los viriones de picornavirus varía de 8 x 10 6 – 9 x 10 6 con una tasa de sedimentación (S 20w) de 140-165S (para partículas vacías S 20w es 70-80S). Su densidad de flotación en CsCl es 1.33-1.45 g cm -3 , dependiendo del género.
Algunas especies son inestables por debajo de pH 7; muchos son menos estables con una fuerza iónica baja que con una fuerza iónica alta. Los viriones son insensibles al éter, cloroformo o detergentes no iónicos.
Los virus son inactivados por la luz cuando se cultivan con, o en presencia de tintes fotodinámicos como el rojo neutro o proflavina. La estabilidad térmica varía con los virus al igual que la estabilización por cationes divalentes.
Ácido nucleico
Los viriones contienen una molécula de ssRNA de sentido positivo, con un tamaño de 6.7-10.1 kb, y que poseen un único ORF largo; Picodicistrovirus caninos (género Dicipivirus), sin embargo, muestran dos ORF, la traducción de cada uno está dirigida por un sitio de entrada ribosomal interno (IRES) separado.
Una cola poli, de longitud heterogénea, se ubica después de la secuencia heteropolimérica 3′-terminal. Una proteína pequeña, VPg (c. 2.2 a 3.9 kDa), está unida covalentemente al extremo 5′.
Las regiones no traducidas (UTR) en ambos extremos contienen regiones de estructura secundaria que son esenciales para la función del genoma.
El 5′-UTR largo (500-1500 nt) incluye un dominio 5′-terminal involucrado en la replicación (por ejemplo, el «virus de la hoja de trébol» del poliovirus) y un IRES de 220-450 nt corriente arriba del sitio de inicio de la traducción.
La mayoría de los elementos IRES picornavirales se pueden asignar a uno de varios tipos (I a V, IGR-IRES), de acuerdo con su estructura secundaria. Entre el dominio 5′-terminal y el IRES puede haber uno o más, pseudonudos.
Un tracto poli se encuentra en la 5′-UTR de los virus de la fiebre aftosa, los virus de la encefalomiocarditis y posiblemente los teschovirus porcinos. La 3′-UTR, que también puede contener un pseudoknot, varía entre 25 y casi 800 nt de longitud.
Proteínas
Además de los principales PC, 1A, 1B, 1C y 1D, y 3B (VPg), descritos anteriormente, las pequeñas cantidades de 1AB (VP0) se ven comúnmente en lugar de una o más copias de 1A y 1B.
La proteína 1A es pequeña en los hepatovirus, y la 1AB no se escinde entre los miembros de Ampivirus, Aquamavirus, Avihepatovirus, Avisivirus, Gallivirus, Kobuvirus, Kunsagivirus, Limnipivirus, Mnipivirus, Virus de Virus Sicinivirusgéneros, y una serie de picornavirus sin clasificar.
Las proteínas ortólogas 1B, 1C, 1D, 2C, 3C y 3D se conservan en todos los picornavirus y pueden usarse para alineamientos de secuencias entre virus, mientras que 1A, 2A, 2B, 3A y 3B son altamente divergentes entre los miembros de diferentes géneros de picornavirus.
Las proteinasas virales son las siguientes: 3C pro , una cisteína proteasa de tipo quimotripsina codificada por todos los picornavirus, realiza la mayoría de las escisiones; 2A pro relacionado con 3C pro es responsable de algunas escisiones en enterovirus, y posiblemente sapelovirus y rabovirus.
La L prouna proteína líder es una cisteína proteasa relacionada con la papaína que se libera de la poliproteína en los afhovirus, los erbovirus y posiblemente los mosavirus.
También, el 2A de aftovirus, aquamaviruses, avihepatoviruses, avisiviruses, cardiovirus, cosaviruses, erboviruses, hunniviruses, kunsagiviruses, limnipiviruses, mischiviruses, mosaviruses, miembros de la especie Parechovirus B, C y D , pasiviruses, potamipiviruses, rosaviruses, senecaviruses, teschovirus.
Lípidos
Algunos picornavirus llevan una molécula similar a la esfingosina («factor de bolsillo») en una cavidad («bolsillo») ubicada dentro de 1D.
La proteína 1A, cuando está presente, generalmente tiene una molécula de ácido mirístico unida covalentemente a la glicina amino terminal. No se encuentra ninguna señal de miristoilación en la secuencia N-terminal VP4 y VP0 de avihepatovirus, dicipivirus, kobuvirus, parechovirus y tremovirus.
Carbohidratos
Ninguna de las proteínas virales está glicosilada.
Organización del genoma y replicación
El ARN virión es infeccioso (Colter et al., 1957, Alexander et al., 1958) y sirve como genoma y como ARNm viral. El inicio de la síntesis de proteínas es estimulado por el IRES.
La traducción del único ORF produce el precursor de la poliproteína (216-277 kDa) a las proteínas estructurales (derivadas de la región P1 del genoma) y las proteínas no estructurales (de las regiones P2 y P3) En muchos virus, P1 está precedido por una proteína líder (L).
En los dicipivirus, ORF1 codifica el precursor de las proteínas estructurales (correspondiente a la región P1), mientras que ORF2 codifica el precursor de la proteína funcional (regiones P2 y P3).
La poliproteína se escinde en proteínas funcionales por proteinasas codificadas por virus. Los intermedios se indican mediante combinaciones de letras (por ejemplo, 3CD, el precursor no escindido de 3C y 3D). Los picornavirus exhiben una organización modular del genoma.
Se cree que ciertos elementos genéticos de las regiones 5′- y 3′-UTR y gen se han intercambiado entre los géneros.
Puede haber una o más regiones genéticas que codifican varias proteínas 2A.
Al lado de 2A pro y el naftp 2A similar a afftovirus, 2A puede tener motivos de secuencia H-box / NC [avihepatovirus, avisivirus, gallivirus, kobuvirus, megrivirus, parechovirus, paserivirus, potamipivirus, sakobuvirus, saliviruses, sicinivirus, tremovirus y varios picornavirus no clasificados ( Hughes y Stanway, 2000).
Puede tambien existir una similitud con el dominio de unión a guanina de tipo AIG1, una NTPasa P-loop con el motivo GxxGxGKS (avihepatovirus, avisivirus); su función en la replicación de virus no está clara.
Las regiones de los genes 2B y 3A varían enormemente entre los géneros, aunque funcionalmente pueden ser homólogas.
Algunos intermedios son estables y cumplen funciones distintas de las de sus productos de escisión (por ejemplo, escisión del poliovirus P1 por 3CD pro , no por 3C pro). Donde ocurre, se piensa que la escisión de 1AB, que acompaña a la encapsidación del ARN, es autocatalítica, pero se desconoce el mecanismo preciso.
Un diseño típico del genoma de picornavirus se puede representar mediante lo siguiente:
- VPg + 5’UTR [1A-1B-1C-1D / 2A-2B-2C / 3A-3B-3C-3D] 3’UTR-poli (A).
Donde “[” y “]” definen la extensión de la región codificante de la poliproteína, “/” representa divisiones primarias y “-” representa las divisiones finales.
Cuando un polipéptido particular está presente solo en algunos miembros del género, se puede mostrar entre paréntesis.
Este esquema también se puede usar para indicar algunas funciones de proteínas o motivos de aminoácidos donde difieren entre los virus (por ejemplo, 2A pro o 2A npgp o 2A H-box / NC o 2A NTPase ).
Puede haber múltiples copias de una región genómica particular en el genoma del picornavirus, incluidas copias repetidas de una región particular (por ejemplo, tres 3B en el genoma del virus de la fiebre aftosa).
También pueden ser diferentes tipos de una región particular (por ejemplo, dos motivos 2A diferentes en el virus Ljungan del género Parechovirus) y tres motivos 2A diferentes en el genoma del virus de la hepatitis A de pato del género Avihepatovirus).
La replicación del ARN viral ocurre en estrecha asociación con estructuras membranosas citoplásmicas reorganizadas.
Estos complejos denominados orgánulos de replicación contienen proteínas derivadas de la totalidad de la región 2BC-P3 de la poliproteína, incluida la polimerasa (pol 3D , una enzima que alarga la cadena del ARN), y 2C (una ATPasa que contiene un motivo de secuencia de unión a nucleótidos).
Se ha demostrado que el componente 3C pro de poliovirus y coxsackievirus se requiere para unirse a la hoja de trébol de ARN 5′-terminal.
La pequeña proteína codificada por el virus, VPg, actúa como un cebador de transcripción para la síntesis de ARN de hebra tanto positiva como negativa.
Antes de la transcripción, dos residuos de uridina se unen covalentemente a la tirosina conservada en la posición 3 en VPg para formar VPgpUpU OH a través de un mecanismo de plantilla que involucra un elemento de replicación que actúa en cis ( cre ) y la polimerasa 3D del virus.
El cre es un bucle del vástago que contiene la secuencia «AAAC» en el bucle y se encuentra en varios lugares del genoma dependiendo de la especie / género de virus. Se han descrito muchos compuestos que inhiben específicamente la replicación.
Mutantes resistentes a, o dependientes de, drogas han sido reportados. Se produce recombinación genética, complementación y mezcla fenotípica.
Las partículas defectuosas, que contienen deleciones en las PC o L, se han producido experimentalmente pero no se han observado en poblaciones de virus naturales.
Antigenicidad
Los serotipos se clasifican, según el género, por protección cruzada, neutralización de la infectividad, fijación del complemento, ELISA específico mediante un formato de captura o inmunodifusión.
Algunos serotipos pueden identificarse mediante la inhibición de la hemaglutinación.
Los serotipos se han determinado para la mayoría de los miembros de los enterovirus, afftovirus, cardiovasculares, erbovirus y teschovirus, pero se reemplazan cada vez más por genotipos (comúnmente denominados «tipos») en la práctica clínica o diagnóstica.
Los géneros no están relacionados antigénicamente cuando se investigan.
Biología
La mayoría de los picornavirus para los cuales se han identificado los hospedadores naturales son específicos para uno o muy pocas especies hospedadoras [las excepciones son el virus de la fiebre aftosa (virus de la fiebre aftosa) y el virus de la encefalomiocarditis (EMCV)].
Los miembros de la mayoría de las especies se pueden cultivar en cultivos celulares. Las células hospedadoras resistentes (p. Ej., Células de ratón en el caso de los poliovirus específicos de primates) a menudo se pueden infectar (por una sola ronda) mediante transfección con ARN desnudo e infeccioso.
La transmisión es horizontal, principalmente por vía fecal-oral, fomite o aerotransportada. La transmisión por vectores artrópodos no se conoce, aunque el EMCV se ha aislado de mosquitos y garrapatas y poliovirus de moscas; por lo tanto, la transmisión mecánica puede ser posible.
La infección es generalmente citolítica, pero las infecciones persistentes son comunes con algunas especies y se informan con otras.
Las células infectadas con poliovirus experimentan una extensa vacuolación a medida que las membranas se reorganizan en complejos de replicación viral.
La infección puede ir acompañada de una rápida inhibición de la traducción dependiente de la tapa de los ARNm celulares (2A pro de poliovirus y L pro de aftovirus son inhibidores poderosos), la síntesis de ARNm y la vía secretora celular (se han implicado poliovirus 2B y 3A).
Criterios de demarcación de género
Una especie de picornavirus es una clase de virus relacionados filogenéticamente que normalmente se espera que compartan (i) un grado significativo de identidad de aminoácidos de las proteínas P1, 2C, 3C y 3D, (ii) monofilia en árboles filogenéticos, (iii) esencialmente idéntica mapas genómicos.
Y finalmente (iv) un grado significativo de compatibilidad en el procesamiento proteolítico, la replicación, la encapsidación y la recombinación genética.
Normalmente se esperaría que los miembros de un género tuvieran estructuras y proteínas IRES homólogas; sus secuencias se agrupan en la misma rama en árboles filogenéticos (monofilia).
Los miembros de todos los géneros de picornavirus conocidos difieren en (i) mapas genómicos que muestran características distintivas en comparación con sus parientes más cercanos.
Seguidamente (ii) divergencia significativa (número de diferencias por sitio entre secuencias) de las proteínas ortólogas que exceden el 66% de P1 cap y 64% de 2C hel, 3C pro y 3D pol [estos valores se basan en los datos de secuencia actuales y pueden variar con los datos adicionales disponibles en el futuro].
Se manifiesta entonces, (iii) falta de homología detectable de proteínas L (si está presente), 2B, 3A, 3B. Si estas reglas no se aplican, se puede proponer una nueva especie o género.
El Grupo de Estudio Picornaviridae recomienda que las designaciones de cepas incluyan información sobre las especies hospedadoras, el número de identificación específico del laboratorio, el país de muestreo, el año de muestreo, por ejemplo, el sello / ABC1234 / USA / 2015.
Cuando se propone una nueva especie de picornavirus, se debe proporcionar un nombre de virus preferido .
Derivación de nombres
Picorna: un acrónimo de p V igilanciadela, i nsensitivity a éter, c oxsackievirus, o virus rphan, r hinovirus, r ibo n ucleic un cid; también, del prefijo » pico » que designa una unidad de medida muy pequeña (equivalente a 10 -12 ) y ARN para designar virus de ARN muy pequeños.
Relaciones filogenéticas
Los virus de cada género de picornavirus son filogenéticamente distintos de los miembros de otros géneros en aquellas regiones del genoma que son ortólogas, es decir, P1 cap, 2C hel , 3C pro y 3D pol.
La divergencia entre los miembros de un género puede ser tan alta como 67% para la poliproteína P1 y 64% para las proteínas 2C, 3C y 3D. La divergencia entre los miembros de diferentes géneros suele ser mayor.
Similitud con otros taxones
La presencia de 1-3 dominios con similitud detectable con la superfamilia de tipo rhv con plegamiento característico («barril-b») en las proteínas de la cápside es común a la mayoría de los miembros del orden Picornavirales .
Asimismo, en todos los miembros de Picornavirales también está presente un «bloque de replicación» que comprende un dominio de ARN-helicasa (P-bucle NTPasa), una peptidasa C-proteinasa y un ARN dependiente de ARN polimerasa 1 (RT-como la superfamilia).y muchos virus parecidos a picorna no clasificados.
Las proteínas de la cápsida y las proteínas del bloque de replicación muestran diversos grados de similitud. La presencia de una pequeña proteína ligada al genoma que también es el cebador de replicación también es común a muchos pequeños virus de ARN de cadena positiva.
Una organización del genoma con un solo marco de lectura abierto y una región del gen que codifica CP en el extremo 5 ‘y la región del gen que codifica las proteínas del bloque de replicación en el extremo 3’ es común a los miembros de Picornaviridae e Iflaviridae.
Patogenicidad
Los picornavirus son un grupo diverso de patógenos virales humanos que, en conjunto, constituyen las causas más comunes de infecciones en humanos en el mundo desarrollado.
Dentro de la familia del picornavirus hay tres grupos conocidos de patógenos humanos: los rinovirus humanos (HRV), los enterovirus (EV) (incluidos los poliovirus, coxsackievirus y echovirus) y los hepatovirus (incluida la hepatitis A).
Los HRV incluyen más de 100 serotipos en dos grupos principales basados en sus receptores celulares. El principal factor de riesgo para la infección por HRV parece ser el contacto con niños pequeños.
Enfermedades que son causadas por el picornaviridae
Los miembros de picornaviridae pueden causar una variedad de enfermedades significativas y muy diferentes, como parálisis, enfermedad de manos, pies y boca, resfriado común, miocarditis y hepatitis, entre otras.
Síntomas
La mayoría de las infecciones por EV sintomáticas en los Estados Unidos se caracterizan por enfermedades leves de EV asociadas con fiebre y síntomas constitucionales, con o sin erupciones cutáneas.
Estas enfermedades son de importancia clínica porque pueden imitar otras enfermedades como la sepsis bacteriana, otras enfermedades virales exantemáticas y las infecciones por herpes simple.
Una revisión reciente de las enfermedades respiratorias asociadas a EV encontró que el 46% de los casos se presentaron con infecciones respiratorias superiores, el 13% con dificultad respiratoria / apnea, el 13% con neumonía, el 12% con otitis media y menos casos de bronquiolitis, sibilancias, crup, y faringoamigdalitis.
Diagnosis
Los nuevos desarrollos en el diagnóstico rápido y la terapia de estas infecciones prometen reducir significativamente la carga de la enfermedad y los costos asociados para las personas afectadas y para la sociedad.
El diagnóstico depende principalmente de aislar y tipear el virus de un hisopo de garganta o una muestra de heces. El análisis del ARN viral en el LCR (por RT-PCR) puede ser una forma rápida y confiable de definir un Picornavirus como la causa de la meningitis.
El valor del análisis serológico es controvertido. Existe un ensayo de fijación del complemento basado en un cóctel de anticuerpos anti-Picornavirus, pero esta prueba carece de sensibilidad.
El diagnóstico depende de una seroconversión documentada o un título de más de 80. Los anticuerpos pueden detectarse mediante una técnica de neutralización basada en un número limitado de serotipos, es decir, los que se encuentran con más frecuencia en infecciones humanas (Coxsackie A7, A9, A16 y B1 a B6).
Los anticuerpos neutralizantes persisten mucho después de la resolución de la infección y, además, pueden ser estimulados por otros antígenos, no necesariamente relacionados.
La seroconversión es una buena evidencia, pero la simple detección de títulos altos de uno o más anticuerpos no es concluyente de ninguna manera y debe interpretarse en el contexto de la naturaleza de los síntomas y cuando aparecieron por primera vez.
Tratamiento
Actualmente, existen pocos tratamientos para muchos de los virus en Picornaviridae. Para los rinovirus, no existen tratamientos aprobados, aunque el ruprintrivir parece prometedor en ensayos clínicos y los éteres de piridazinil oxima pueden resultar útiles.
Los tratamientos contra el poliovirus son necesarios para complementar el plan de erradicación de la poliomielitis de la Organización Mundial de la Salud para tratar las infecciones causadas por la reversión del virus de la vacuna atenuada y para complementar el control de la cobertura de la vacuna en las áreas endémicas de la poliomielitis.
Sin embargo, no se han desarrollado compuestos prometedores para el tratamiento del poliovirus debido a la eficacia de las vacunas en uso.
Los inhibidores de amplio espectro desarrollados para otros picornavirus pueden ser útiles para las infecciones por poliovirus.
Las infecciones por coxsackievirus en niños y bebés están siendo tratadas con pleconaril con cierta eficacia para reducir la mortalidad y mejorar la recuperación, aunque el tratamiento a menudo es sobre una base de uso compasivo.
No hay terapias para las infecciones por echovirus.
Se está realizando muy poca investigación de descubrimiento de fármacos para desarrollar inhibidores de las infecciones por ecovirus, probablemente debido a la inhibición de amplio espectro que presentan los agentes de unión a la cápside y los inhibidores de la proteasa descubiertos para el tratamiento de otros picornavirus.
Por ejemplo, los éteres de piridazinil oxima son inhibidores de la mayoría de los echovirus. Los tratamientos para las infecciones por enterovirus también son limitados, aunque en un ensayo clínico pequeño, La milrinona pareció reducir la mortalidad y mejorar la recuperación del edema pulmonar inducido por EV71.
Por lo tanto, estos resultados enfatizan fuertemente la necesidad del desarrollo de compuestos potentes y no tóxicos para el tratamiento de infecciones por picornavirus.